

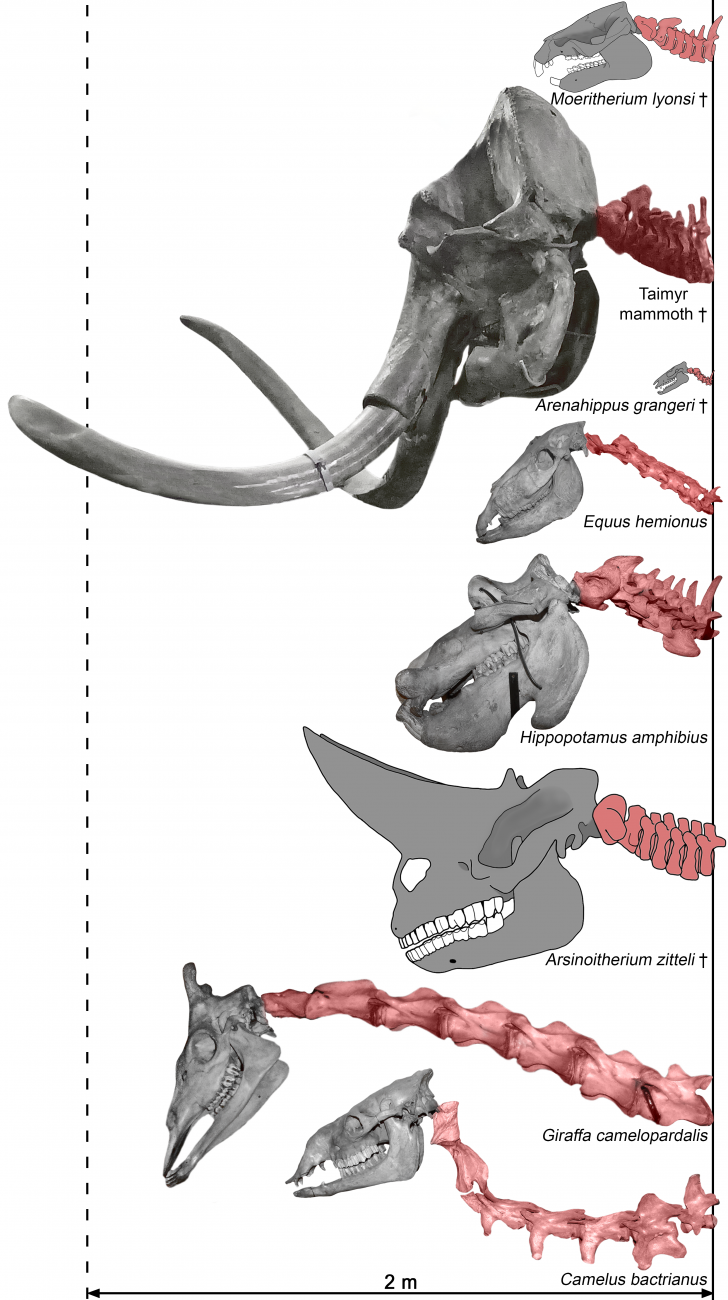
In a new article published in the Journal of Anatomy, Ruslan Belyaev and Natalia Prilepskaya, employees of the Laboratory of Ecology, Physiology, and Functional Morphology of Higher Vertebrates at the Institute of Ecology and Evolution of the Russian Academy of Sciences, together with colleagues from Yakutia, Israel, and Belgium, conducted a large-scale study of body proportions in large herbivorous mammals. The work examined dozens of skeletons of modern elephants, even-toed and odd-toed ungulates, as well as some fossil representatives of these orders, including woolly and steppe mammoths, the American mastodon, megabelodon, hyrachyus, etc. Comparison of modern odd-toed ungulates (horses, rhinoceroses, tapirs) and elephants with their Paleogene ancestors made it possible to demonstrate how the body plan changed in these phylogenetic lines over tens of millions of years of evolution, and to offer biomechanical interpretations of the observed changes.
The Cenozoic is rightfully considered the era of mammals. After the Cretaceous-Paleogene extinction, mammals became full-fledged masters of the planet, occupying many ecological niches previously inaccessible to them. In many phylogenetic lines, large and gigantic forms appeared, some of which exist today. The appearance of large herbivorous mammals is extremely diverse, the differences in the body proportions of elephants and giraffes, horses and rhinoceroses, bison and hippos make their exterior instantly recognizable to our eyes. However, in addition to the striking differences, these animals also have numerous similarities in the biomechanics of their bodies, which were formed in response to the adaptation of the musculoskeletal system to gigantism.
The earliest known ancestors of proboscideans and odd-toed ungulates were quite small forms. Thus, the oldest known proboscidean – Eritherium azzouzorum from the Paleocene (~60 million years) of Morocco – was slightly larger than the hyrax and weighed 5-6 kg. The fairly well-known plesiomorphic odd-toed ungulates from the early Eocene (50-55 million years) of North America and Eurasia were the size of a dog and weighed from 9-10 to 20-50 kg. How have modern elephants, horses and rhinoceroses changed compared to their Paleogene ancestors, apart from an impressive increase in size? In both orders, changes occurred in the course of evolution associated with both allometric growth and deep specialization of the locomotor apparatus. Moreover, if allometric changes turn out to be, in many ways, similar to each other, then the directions of specialization of the locomotor apparatus in elephants and ungulates turn out to be directly opposite.
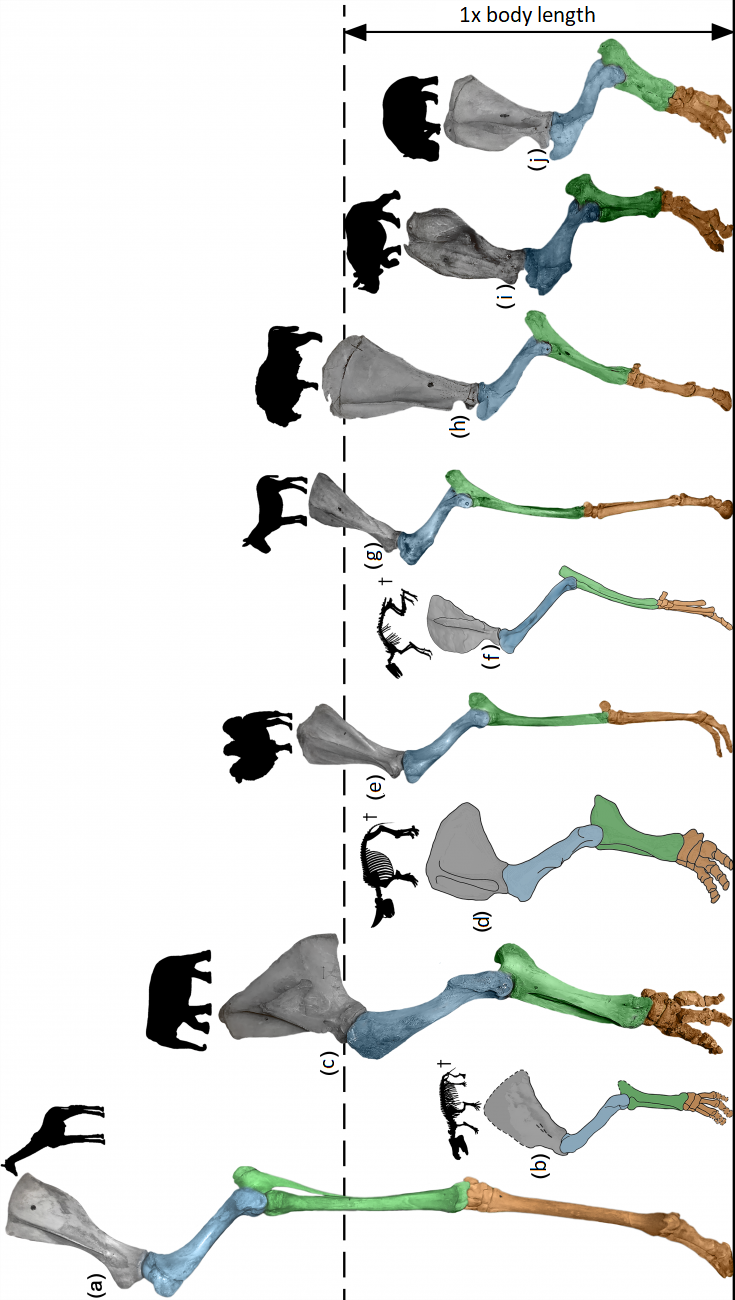
Figure 2. Relative height of the forelimb in large herbivorous mammals: (a) giraffe, (b) moeritherium (late Eocene of Africa), (c) Indian elephant, (d) arsinoitherium (late Eocene of Africa), (e) two-humped camel, (f) arenahippus (early Eocene of North America), (g) kulan, (h) bison, (i) Indian rhinoceros, (j) hippopotamus.
In the case of elephants, the musculoskeletal system specializes in an extremely economical mode of locomotion based on the inverse pendulum principle. Surprisingly, this principle is also implemented in human bipedal locomotion. In the course of evolution, proboscideans lose not only the ability to jump and use asymmetric gaits, including gallop, but also the ability to run as such. Even during the fastest locomotion, one of the elephant's limbs always maintains support on the substrate. Along with the loss of the ability to use a gallop, proboscideans almost completely lost the ability of the back to bend in the sagittal plane, their hind limbs lost their three-link structure (thigh, shin, foot) becoming two-link, the joints of the limbs straightened, which made the leg columnar (Figure 2). Such a columnar limb turns into a kind of pole, allowing for a highly efficient transition of the kinetic energy of the moving center of mass into potential and vice versa. At the same time, energy during locomotion is practically not spent on vertical movements of the center of mass, which in elephants during fast movement is 7 times lower than, for example, in a tapir. As a result, elephant walking using the inverted pendulum principle is one of the most energy-efficient ways of movement among terrestrial vertebrates.
During evolution, the increase in the body size of proboscideans is accompanied by a significant increase in the height of their forelimbs and hindlimbs. Thus, compared to the Moeritherium from the late Eocene of Africa (El Fayoum, Egypt), the height of the limb of modern elephants relative to their own body length increases more than twice (Figure 2b, c). This allows them to increase the stride length (and, as a consequence, the speed of movement) and simultaneously reduce vertical oscillations of the center of mass during locomotion. The elongation of the limbs occurs due to the length of their proximal segments (scapula, shoulder and forearm in the forelimb, thigh and lower leg in the hind limb), while the contribution of the hand and foot to the limb length becomes minimal. Long limbs were also characteristic of some groups of extinct proboscideans, including mastodons and amebelodons. However, the longest limbs among the studied proboscideans were possessed by mammoths – Mammuthus primigenius and M. trogontherii. The taller pendulums of the limbs would have provided even more energy-efficient locomotion than modern elephants, making the limbs ideal for long seasonal migrations and overcoming land and water obstacles.
In ungulate mammals, the exact opposite optimization of the locomotor apparatus occurs. If proboscideans lose the ability to run, then ungulates, on the contrary, become one of the most specialized in running. They retain the three-link structure of the limbs, characteristic of plesiomorphic therian mammals, and their joints remain visually “bent” when compared with the practically straight limbs of elephants. Three-link limbs, the joints of which are oriented in a zigzag manner, allow ungulates to optimize the kinematics of the joints and reduce energy costs for undesirable mechanical work of muscles directed against each other inside the limb. This significantly reduces energy costs when running, especially with the use of asymmetric gaits.
During the evolution of odd-toed ungulates, there is a significant reorganization of the locomotor apparatus, which is most obvious in horses. The relative height of the limbs in these animals becomes significantly greater than that of their small Paleogene ancestors (Figure 2e, g). Moreover, unlike elephants, the increase in height occurs primarily due to the lengthening of the distal segments, i.e. the hand and foot. Their length in equines increases almost twice as much as in their Paleogene ancestors. Such elongation of the limbs allows the bulk of the muscles to be shifted proximally, making the lower half of the leg relatively lighter. This reduces the expenditure of mechanical energy, reduces the moment of inertia of the limb and increases the speed of its transfer during running. Along with the increase in the absolute and relative length of the limbs, equines experience a twofold increase in the relative length of the neck and a one and a half fold increase in the relative length of the skull. A significantly longer neck and head allow horses to retain the ability to graze at the level of the substrate despite the significantly increased height of the legs.
The work is published in the journal: Ruslan I. Belyaev, Gennady G. Boeskorov, Alexander N. Kuznetsov, Mathys Rotonda, Natalya E. Prilepskaya. 2024. Comparative study of the body proportions in Elephantidae and other large herbivorous mammals. Journal of Anatomy.