

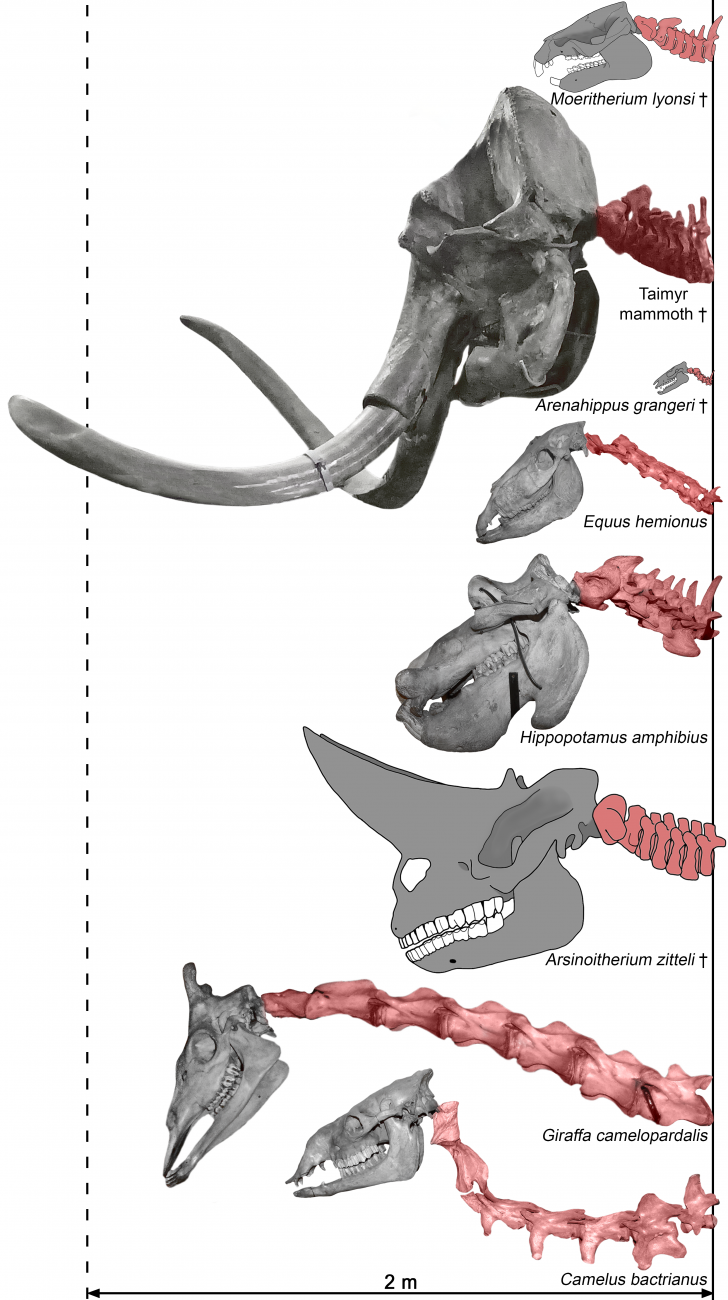
Сотрудники Лаборатории экологии, физиологии и функциональной морфологии высших позвоночных Института проблем экологии и эволюции им. А.Н. Северцова РАН Руслан Беляев и Наталья Прилепская вместе с коллегами из ИГАБМ СО РАН (Якутия), Technion (Израиль) и RMCA (Бельгия) провели масштабное исследование пропорций тела у крупных растительноядных млекопитающих.. В проведенной работе были изучены десятки скелетов современных слонов, парно- и непарнокопытных, а также некоторые ископаемые представители этих отрядов, включая шерстистого и степного мамонтов, американского мастодонта, мегабелодона, гирахиуса и пр. Сравнение современных непарнокопытных (лошадей, носорогов, тапиров) и слонов с их палеогеновыми предками позволило продемонстрировать, как изменился план строения тела в этих филогенетических линиях за десятки миллионов лет эволюции, а также предложить биомеханические интерпретации наблюдаемым изменениям.
Кайнозой по праву считается эрой млекопитающих. После мел-палеогенового вымирания млекопитающие стали полноценными хозяевами планеты, заняв многие ранее недоступные для них экологические ниши. Во многих филогенетических линиях появились крупные и гигантские формы, некоторые из которых существуют и сегодня. Внешний вид крупных растительноядных млекопитающих крайне разнообразен, различия в пропорциях тела слонов и жирафов, лошадей и носорогов, бизонов и бегемотов делает их экстерьер мгновенно узнаваемым для наших глаз. Однако, помимо бросающихся в глаза различий эти животные обладают и многочисленными сходствами биомеханики их тел, которые сформированы в ответ на адаптацию опорно-двигательного аппарата к гигантизму.
Наиболее ранние известные предки хоботных и непарнокопытных были достаточно мелкими формами. Так древнейшее известное хоботное – эритерий (Eritherium azzouzorum) из палеоцена (~60 млн лет) Марокко – немного превосходил размерами дамана и весил 5-6 кг. Достаточно хорошо известные плезиоморфные непарнокопытные из раннего эоцена (50-55 млн лет) Северной Америки и Евразии были размером с собаку и весили от 9-10 до 20-50 кг. Как же изменились современные слоны, лошади и носороги по сравнению со своими палеогеновыми предками помимо впечатляющего увеличения в размерах? В обоих отрядах в ходе эволюции происходили изменения, связанные как с аллометрическим ростом, так и с глубокой специализацией локомоторного аппарата. Причем, если аллометрические изменения оказываются, во-многом, схожими между собой, то направления специализации локомоторного аппарата у слонов и копытных оказываются прямо противоположными.
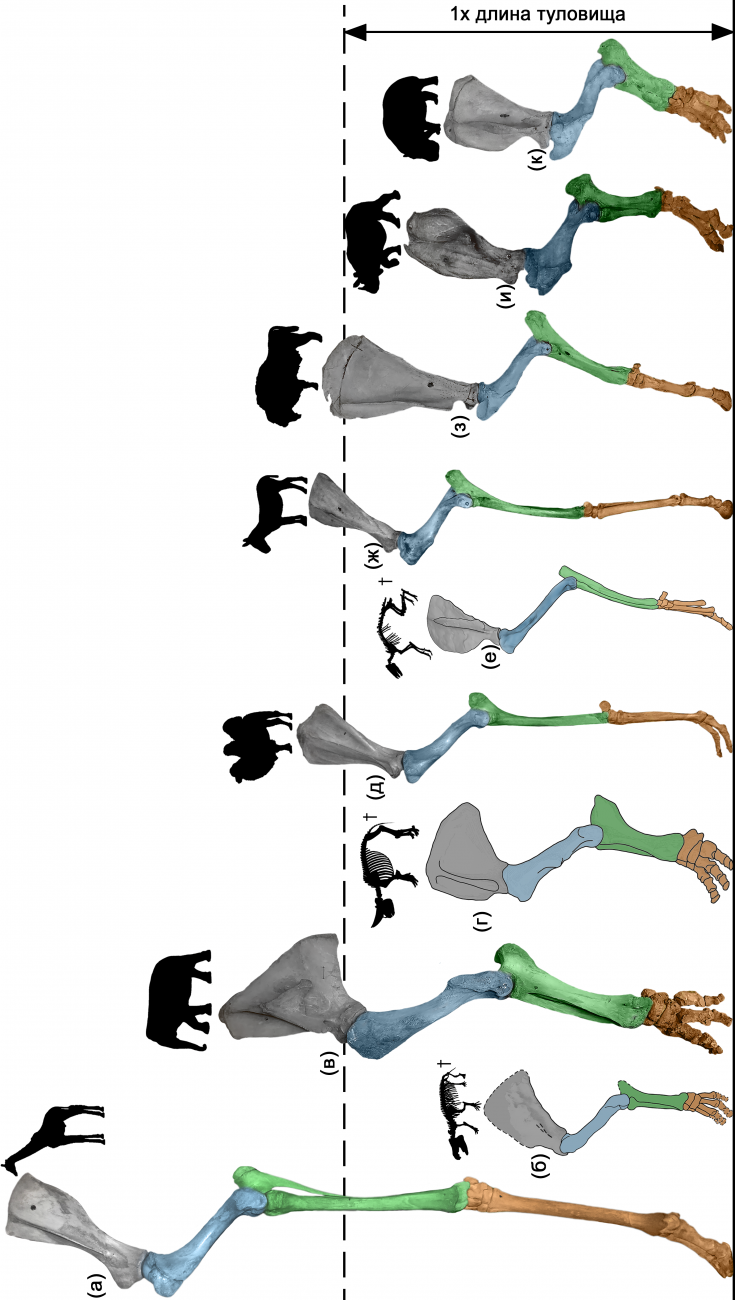
Рисунок 2. Относительная высота передней конечности у крупных растительноядных млекопитающих: (а) жираф, (б) меритерий (поздний эоцен Африки), (в) индийский слон, (г) арсинойтерий (поздний эоцен Африки), (д) двугорбый верблюд, (е) аренагиппус (ранний эоцен Северной Америки), (ж) кулан, (з) зубр, (и) индийский носорог, (к) гиппопотам.
В случае слонов происходит специализация опорно-двигательного аппарата к крайне экономичному способу передвижения, основанному на принципе обратного маятника. Удивительным образом, именно этот принцип реализован и в двуногой локомоции человека. В ходе эволюции хоботные теряют не только способность к прыжкам и использованию ассимитричных аллюров, включая галоп, но и способность к бегу как таковому. Даже при самой быстрой локомоции одна из конечностей слона всегда сохраняет опору на субстрат. Вместе с потерей способности использовать галоп, хоботные практически полностью утратили способность спины к сгибанию в сагиттальной плоскости, их задние конечности утратили трехзвенную структуру (бедро, голень, стопа) став двухзвенными, суставы конечностей выпрямились, что сделало ногу колоннообразной (Рисунок 2). Такая колоннообразная конечность превращается в своеобразный шест, позволяющий осуществлять высокоэффективный переход кинетической энергии движущегося центра масс в потенциальную и наоборот. При этом, энергия во время локомоции практически не тратится на вертикальные передвижения центра масс, которые у слонов во время быстрого передвижения в 7 раз ниже, чем, например, у тапира. В результате, ходьба слонов с использованием принципа перевернутого маятника является одним из наиболее энергоэффективных способов передвижения среди наземных позвоночных.
В ходе эволюции увеличение размеров тела хоботных сопровождается существенным увеличением высоты их передних и задних конечностей. Так, по сравнению с меритерием из позднего эоцена Африки (Эль-Файюм, Египет), высота конечности современных слонов относительно длины их собственного тела увеличивается более чем вдвое (Рисунок 2б, в). Это позволяет им увеличить длину шага (а как следствие и скорость передвижения) и одновременно сократить вертикальные колебаний центра масс при локомоции. Удлинение конечностей происходит за счет длины их проксимальных сегментов (лопатка, плечо и предплечье в передней конечности, бедро и голень в задней), в то время как вклад кисти и стопы в длину конечности становится минимальным. Высокие конечности были характерны и для некоторых групп вымерших хоботных, включая мастодонтов и амебелодонов. Однако наиболее высокими конечностями среди изученных хоботных обладали мамонты – Mammuthus primigenius и M. trogontherii. Более высокие маятники конечностей должны были обеспечивать еще более энергоэффективное передвижение, чем у современных слонов. Это делало конечности идеально подходящими для совершения длинных сезонных миграций и преодоления наземных и водных препятствий.
У копытных млекопитающих происходит прямо противоположная оптимизация локомоторного аппарата. Если хоботные теряют способность к бегу, то копытные, напротив, становятся одними из наиболее специализированных к бегу животных. Они сохраняют трехзвенную структуру конечностей, характерную для плезиоморфных териевых млекопитающих, а их суставы остаются визуально “подсогнутыми” при сравнении с практически прямыми конечностями слонов. Трехзвенные конечности, суставы которых зигзагообразно ориентированы, позволяют копытным оптимизировать кинематику суставов и снизить энергетические затраты на нежелательную механическую работу мускулов, направленную друг против друга внутри конечности. Это существенно снижает энергетические затраты при беге, особенно с использованием ассимитричных аллюров.
В ходе эволюции непарнокопытных происходит существенная перестройка локомоторного аппарата, которая наиболее очевидна у лошадей. Относительная высота конечностей у этих животных становится существенно больше, чем у их мелких палеогеновых предков (Рисунок 2е, ж). Причем в отличие от слонов, увеличение высоты происходит в первую очередь за счет удлинения дистальных сегментов, то есть кисти и стопы. Их длина у лошадиных увеличивается по сравнению с палеогеновыми предками практически вдвое. Подобное удлинение конечностей позволяет сместить основную массу мускулатуры проксимально, делает нижнюю половину ноги относительно более легкой. Это позволяет сократить расход механической энергии, уменьшить момент инерции конечности и увеличить скорость ее переноса во время бега. Вместе с увеличением абсолютной и относительной длины конечностей у лошадиных происходит двухкратное увеличение относительной длины шеи и полуторакратное увеличение относительной длины черепа. Существенно более длинная шея и голова позволяют лошадям сохранить возможность пастись на уровне субстрата несмотря на существенно увеличившуюся высоту ног.
Работа опубликована в журнале: Ruslan I. Belyaev, Gennady G. Boeskorov, Alexander N. Kuznetsov, Mathys Rotonda, Natalya E. Prilepskaya. 2024. Comparative study of the body proportions in Elephantidae and other large herbivorous mammals. Journal of Anatomy.