


Умение быстро бегать является базовой адаптацией, помогающей в борьбе за выживание многим наземным животным. Бег позволяет хищным животным активно охотиться, а их жертвам убегать от опасности. Во время бега важную роль играют как движения конечностей, так и позвоночного столба. Исходный для наземных тетрапод способ локомоции был унаследован ими от рыбообразных предков. Выйдя на сушу, они сохранили тот же паттерн движений спины, который характерен для плавания рыб, — боковые изгибы тела. При этом у рыб продвижение осуществляется за счет отталкивания от воды всем телом назад и вбок, а у рептилий — за счет отталкивания лапами от земли. Однако в отличие от рептилий, спина млекопитающих при ходьбе и беге практически не изгибается вбок. Вместо этого она способна гнуться вертикально, то есть вверх-вниз. Это важнейшее свойство позволило наземным млекопитающим освоить галоп — скоростной и очень экономичный способ передвижения, а водным млекопитающим, таким как киты и сирены, плавать с помощью вертикальных (а не боковых, как у рыб, амфибий и рептилий) движений хвоста.
Несмотря на фундаментальную роль подвижности позвоночника, изученность этой темы остается крайне фрагментарной. Так среди современных непарнокопытных фактические данные о гибкости спины доступны только для домашней лошади. В своем новом исследование специалисты из ИПЭЭ РАН и ПИН РАН использовали разработанную ими раннее математическую модель межпозвонкового сустава, откалиброванную для расчета подвижности в суставах копытных млекопитающих с помощью экспериментальных данных. Эта модель позволяет рассчитывать амплитуду движения в межпозвонковом сочленении, используя размеры суставных поверхностей двух соединяющихся в суставе позвонков. В результате ими были изучены представители пятнадцати из шестнадцати современных видов непарнокопытных, а также впервые была реконструирована подвижность спины у эоценовых предков лошадей, живших примерно 55 миллионов лет назад.
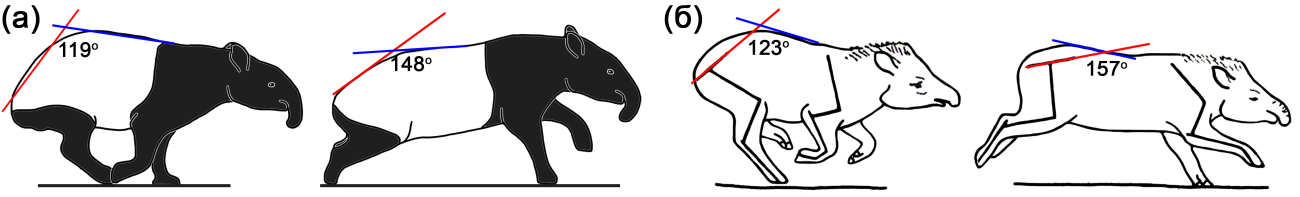
Непарнокопытные – это древняя и в прошлом крайне разнообразная группа млекопитающих. В отличие от парнокопытных (буйволов, антилоп, оленей, свиней, верблюдов и т. п.), расцвет разнообразия которых начался 20–25 миллионов лет назад и значительная часть которых сохранилась до наших дней, основная адаптивная радиация и дивергенция непарнокопытных происходила в раннем эоцене, более 50 миллионов лет назад. Современные непарнокопытные представляют собой лишь скромные остатки некогда обширной и разнообразной группы. Они включают крупных растительноядных млекопитающих, принадлежащих к трем семействам: тапировые (Tapiridae), носороговые (Rhinocerotidae) и лошадиные (Equidae). Их представители довольно сильно отличаются друг от друга и имеют специфический и легко узнаваемый экстерьер. Лошади являются наиболее хорошо изученными животными, использующими для передвижения дорсостабильный галоп. Во время бега галопом их спина лишь незначительно изгибается в вертикальной плоскости (это и делает верховую езду на этих животных настолько комфортной). Как было показано в новом исследовании, вертикальная гибкость поясницы невелика у всех лошадиных — лошадей, зебр и ослов. Она совсем немного превышает минимум, известный для парнокопытных, который характерен для таких огромных животных, как жирафы и бегемоты.
Однако, несмотря на низкую подвижность поясничного отдела у лошадиных, их спина является наиболее гибкой среди современных непарнокопытных. Вертикальная подвижность в поясничной области у тапиров даже ниже, чем у лошадей, и находится на одном уровне с жирафами и бегемотами, а у носорогов — самая низкая среди всех современных парно- и непарнокопытных.
Это ставит закономерный вопрос: почему спина у непарнокопытных настолько малоподвижна? Является ли это состояние для представителей отряда исходным или возникшим вторично? Для ответа на эти вопросы в проведенном исследование была впервые оценена подвижность спины у ископаемого предка непарнокопытных. Объектом для изучения стал эквоид Arenahippus grangeri из раннего эоцена Северной Америки. Подвижность поясничного отдела позвоночника у аренагиппуса оказалась в полтора раза выше, чем у современных лошадиных, и в 2–3 раза выше, чем у рецентных тапиров и носорогов.
Схожая с аренагиппусом морфология была характерна и для других палеогеновых непарнокопытных. Это были мелкие животные, так Arenahippus имел длину около 60–70 см и весил примерно 9 кг, тапироид Heptodon — около 15 кг, а риноцеротоид Hyrachyus — около 20–50 кг. Для них было характерно большее, чем у современных непарнокопытных число поясничных позвонков (семь против трех-четырех у носорогов, четырех-пяти у тапиров, пяти-шести у лошадиных), а также большая относительная длина поясницы (~40% от суммы длин грудного и поясничного отделов). Что особенно сильно отличает их от современных носорогов, у которых относительная длина поясницы короче в три раза (~14%).
Эти особенности указывают на то, что все палеогеновые непарнокопытные активно использовали вертикальную гибкость спины во время галопа. Располагаемая вертикальная гибкость в поясничном отделе у A. grangeri соответствует многим мелким и средним по размеру парнокопытным, использующим для бега прыжковые формы галопа. Авторы работы предложили гипотезу о том, что снижение подвижности спины и выработка дорсостабильного галопа у современных непарнокопытных могут быть связаны с особенностями их пищеварения. Увеличение размеров тела у всех групп непарнокопытных в ходе эволюции очевидным образом сопровождалось увеличением потребности в пище, что привело к формированию таких хорошо изученных черт, как очень высокие коронки зубов (гипсодонтия) и усложнение морфологии жевательной поверхности щечных зубов (лофодонтия). Это же привело и к усложнению пищеварительного тракта.
Пищеварительная система у растительноядных животных характеризуется увеличением длины кишечника и формированием специальных камер для ферментации целлюлозы. У непарнокопытных ферментация целлюлозы происходит в толстом кишечнике. Так, объем желудка у лошади составляет в среднем около 19 литров, в то время как объем одной лишь слепой кишки 26–30 литров, а всех других отделов толстого кишечника — в среднем 76 литров. Вследствие этого заполненный пищей кишечник составляет значительную долю от массы тела, причем этот вес приурочен именно к его задней половине. Вероятно, что чем тяжелее в ходе эволюции становилась пищеварительная система, тем большую нагрузку испытывали поясничные мышцы. Поэтому, чтобы носить тяжелый кишечник, непарнокопытным необходимо было усилить мышцы поясницы. Это можно было сделать двумя способами. Первый — нарастить мышечную массу. Но для этого требовалось бы отобрать ее у каких-то других мышц. Второй способ — уменьшить длину мышечного волокна (то есть амплитуду их сокращения). Это позволило бы пустить все высвободившуюся массу на увеличение количества волокон, то есть силы.
Такое перераспределение между длиной и поперечным сечением в пользу последнего существенно ограничивает вертикальную подвижность поясницы. В результате чего в эволюционной истории лошадиных произошел переход от передвижения с помощью следующих друг за другом амплитудных прыжков (прыжково-скоростного галопа), который был характерен для их палеогеновых предков, к бегу с минимальными вертикальными колебаниями центра тяжести и более высокой частотой шагов (скоростному галопу), характерному для всех современных представителей этого семейства.
Источник: Ruslan I. Belyaev, Alexander N. Kuznetsov, Natalya E. Prilepskaya. Truly dorsostable runners: Vertebral mobility in rhinoceroses, tapirs, and horses // Journal of Anatomy. V. 242. Issue 4. P. 568–591. https://doi.org/10.1111/joa.13799. Ссылка: https://onlinelibrary.wiley.com/doi/10.1111/joa.13799