

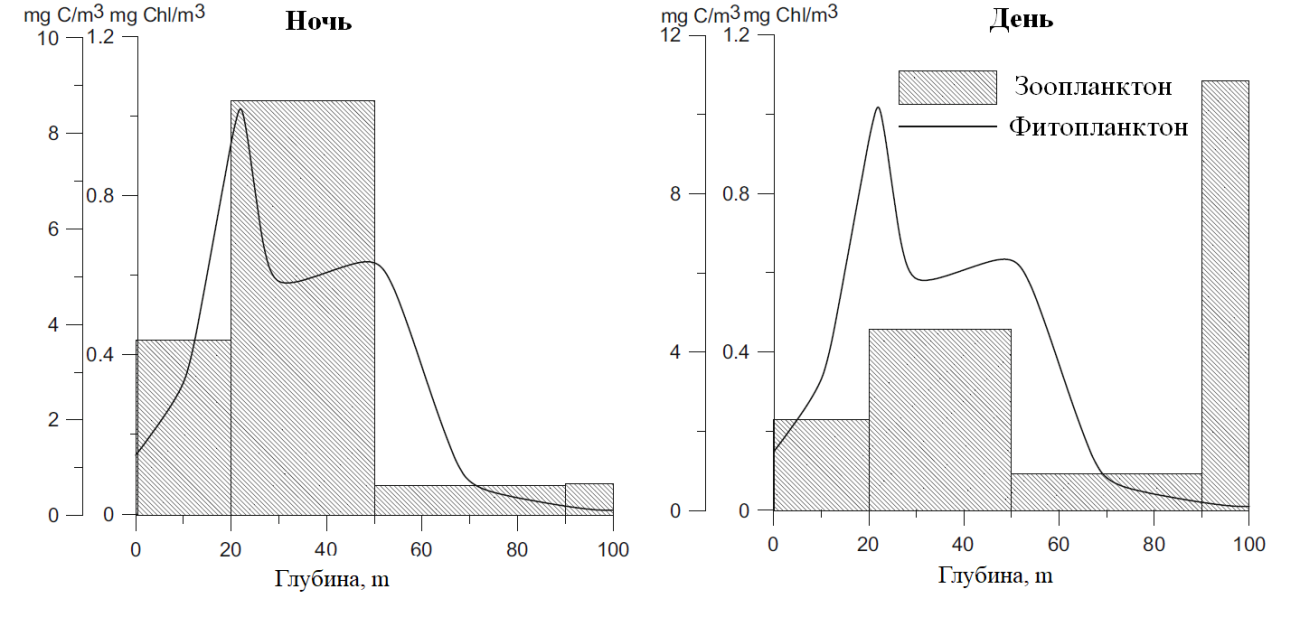
Распределение живых организмов в физическом пространстве часто неоднородно, что описывается в математических моделях пространственно-экологических систем. Изначально математические модели пространственного распределения видов основывались на принципе локальных («точечных») взаимодействий между описываемыми организмами в пространстве. Однако, в настоящее время учет нелокальных взаимодействий приводит к переосмыслению концепций построения математических моделей в экологии. Сценарии возникновения нелокальных взаимодействий весьма многообразны. Например, растения конкурируют за ресурсы (воду) не только со своими непосредственными соседями, но и с другими растениями, произрастающими на обширной территории. Другой важный пример – популяции зоопланктона в водной толще, которые совершают суточные вертикальные миграции. Пример вертикальных распределений зоопланктона в дневное и ночное время показан на рисунке, основанном на натурных наблюдениях: днем зоопланктон остается в глубинных водах, однако на ночь он поднимается в слои воды, богатые фитопланктоном и питается в этих слоях. Даже в течение нескольких часов зоопланктонные организмы могут значительно менять свое вертикальное положение. Поэтому, в математических моделях, оперирующих среднесуточными (или более продолжительными) показателями, для описания подобных популяции необходимо использовать нелокальные, т.е. интегральные методы описания системы.
Несмотря на многочисленные подтверждения важности учета нелокальных взаимодействий, существующие методы моделирования экологических систем по-прежнему основываются на использовании локальных функций. Этот фактор в значительной мере уменьшает предсказательную точность современных математических моделей в пространственной экологии, что тормозит прогресс в данной области.
В своей статье старший научный сотрудник Института проблем экологии и эволюции им. А.Н. Северцова РАН (ИПЭЭ РАН) Андрей Морозов, к.ф.-м. н., подчеркивает важность включения нелокальных взаимодействий в пространственно-временных моделях пищевых цепей. В частности, в работе показано, что учет «интегральных» взаимодействий в моделях хищник-жертва (зоопланктон-фитопланктон) приводит к дополнительной устойчивости системы, которую не предсказывают «классические» модели, основанные на локальных взаимодействиях. Математический аппарат таких моделей включает интегро-дифференциальные уравнения с разными типами ядер в интегральных членах. Однако, использования моделей с нелокальными взаимодействиями так же имеет существенный недостаток: математическая сложность таких моделей значительно возрастает, что приводит к необходимости разработки новых эффективных численных алгоритмов.
“Включение нелокальных взаимодействий так же важно эко-эволюционных моделях, учитывающих, как экологические, так и эволюционные процессы. В этом случае вместо физического пространства взаимодействующих организмов рассматривается пространство фенотипов, описывающееся математически сходными уравнениями с интегральными членами, моделирующими мутации. Интересно, что такой же математический аппарат используется в настоящее время для построения эффективных алгоритмов глобальной оптимизации на основе эволюционных алгоритмов ”, - рассказал А.Ю. Морозов.
Результаты опубликованы в журнале Physics of Life Reviews. Morozov, A., 2026. On implementation of nonlocal terms in models in ecology and evolution. Vol., 56, pp. 208-21.