


Весьма сомнительно, чтобы нашлись еще другие животные,
которые в истории земной коры заняли бы столь видное место.
Ч. Дарвин
Дождевые черви, в отличие от многих других длинных и почти круглых животных, вызывают скорее симпатию, нежели прочие эмоции. С легкой руки Дарвина широко известна выдающаяся роль в повышении плодородия почв дождевых червей, «самоотверженно» пропускающих через свое тело раз за разом чуть ли не весь поверхностный слой почвы парков и газонов в течение всего лишь нескольких лет. Ныне никого не удивишь рекламой «вермикультур» с предложениями купить дождевых червей для плодовых садов, грядок, компостных ям, для цветочных горшков в дом и т.д. Однако мало кто знает, что степень изученности этих замечательных животных несопоставимо ничтожна по сравнению с их ролью и в природе, и в лаборатории — как модельного объекта для разнообразных исследований. К примеру, на всей не затронутой хозяйственной деятельностью равнинной Сибири, как считается, обитает всего несколько видов дождевых червей. Они выживают зимой при лютых морозах в самой верхушке почвы, порой не прикрытой снегом. Завезенные сюда даже самые «лучшие» (из питомников) дождевые черви вымерзнут, не сохранив потомства. А местные — капризны в другом отношении: не желают жить на окультуренных землях, казалось бы, в намного лучших условиях.
Действительно ли, что на этой громадной территории — от Урала до Чукотки — разнообразие наших червей так мало?
Одно лицо на всех
Для непосвященных почти все дождевые черви на одно лицо — их внешний вид разнообразием не отличается. Одни живут в почве под подстилкой и в ней, другие — чуть глубже, третьи — более чем в двухметровых норах, которые сами же устраивают. И жители верхних горизонтов почвы, и норники кормятся в подстилке полуразложившейся или даже нетронутой органикой. Обитатели гумусовых горизонтов заглатывают саму почву, извлекая из нее что могут и насыщая ее микроорганизмами. Впрочем, не будем повторять дарвиновскую схему, старательно и успешно разрабатываемую многими поколениями биологов разных профилей. Заметим лишь, что пищевая специализация не отражается на морфологии яркими чертами, как не сказывается на ней и способ передвижения в плотном субстрате; впрочем, живущие в глубоких норах всегда крупнее обитателей поверхности.
И в верхних, и в нижних горизонтах движение возможно лишь путем раздвигания телом менее или более плотной почвы. Никаких специальных органов для этого у дождевых червей, как и у всех олигохет, нет. У ближайших родственников (прародителей) — многощетинковых червей (Polychaeta) — развиты примитивные конечности. Это так называемые параподии — парные выросты туловищных сегментов, располагающиеся по бокам тела метамерно, на каждом сегменте. У дождевых червей в процессе эволюции они утрачены.
Передвижению в толще земли помогают щетинки, расположенные по бокам сегментов. Они столь малы, что невооруженным глазом не разглядеть, однако благодаря им гладкий влажный червь, взятый в руки, ощущается неожиданно шершавым. Щетинки — консервативный, а потому важный для систематики признак, их расположение на сегменте имеет диагностическое значение. Разнообразие окраски также невелико: пигментация дождевых червей обычно варьирует от ее отсутствия до темно-бурого цвета. Существует мнение, что червь приобретает ее в результате поглощения растительных пигментов из опада. Общий тип среды обитания и одинаковый способ передвижения задали единый, достаточно простой план внешнего строения дождевых червей.
В целом же внешними диагностическими признаками для разделения дождевых червей на виды служат положение (на каком сегменте) пояска (на котором формируется яйцевой кокон) и половых отверстий, форма головной лопасти, расположение («сближенность») щетинок, иногда размер и окраска тела. Зачастую эти признаки имеют значительную внутривидовую изменчивость, что, конечно, затрудняет определение, поэтому используются также внутренние анатомические отличия.
Видовое однообразие
Общее число таксонов дождевых червей, выявляемых по приведенным критериям, невелико. Мировая фауна содержит примерно 3,7 тыс. видов [1], тогда как на территории России известно всего около 50 [2]. Очевидно, что по сравнению с большинством других групп беспозвоночных видовое разнообразие дождевых червей «несправедливо» мало, учитывая приходящуюся на них биомассу в сообществах почвенных животных. Подавляющая их часть — эндемики-горцы (кавказские, уральские, алтайские) или обитатели регионов с особой палеогеографией (например, Приморья). Есть среди них гиганты (свыше метра в длину), правда, не в России. Есть неотразимые по окраске: черные с зеленым или синим перламутовым отливом, встречающиеся только в предгорьях и долинах Сихотэ-Алиня и на Амуре черви Drawida ghilarovi, которые относятся к широко распространенному тропическому семейству Moniligastridae. Но и гиганты, и «красавцы» — безусловное исключение среди похожих друг на друга (для неспециалиста) даже эндемичных видов.
Однообразие внешности дождевых червей сказалось на отсутствии русских названий для подавляющей части видов; бытуют лишь жаргонные обозначения у рыболовов (например, «морячки» — для Eisenia fetida) и огородников.

На всей же равнинной части России, от западной границы до Дальнего Востока, широко распространены лишь около полутора десятков видов. Большинство из них — космополиты, легко и широко расселяющиеся, в том числе и при непреднамеренном участии человека: начиная с колонизации территории Сибири казаками и крестьянами-переселенцами и по сию пору дождевые черви перевозятся с различными грузами. Хороший пример тому — шесть завезенных видов на окультуренных землях вблизи Магадана. Они живут здесь на удалении более 1000 км от своих ареалов, отгороженные непригодными для обитания большинства дождевых червей лиственничными редколесьями. Это Lumbricus rubellus, Dendrobaena octaedra, Dendrodrilus rubidus, Eisenia nordenskioldi pallida, Eisenia fetida, Allolobophora parva. Но от побережья вглубь континента они не проникают; точнее сказать — может быть, их и завозят в поселки с овощами время от времени, но они не удерживаются. Так же ведут себя некоторые виды слизней — потребителей всякой зелени, их находят в свежих партиях овощей, но не на огородах.
Другая часть видов червей равнинных территорий Сибири, напротив, «оседлая», причем нередко в такой мере, что полоска непригодной для них почвы становится непреодолимым препятствием; нужны столетия, чтобы сменилась топография ландшафта, появились коридоры для дальнейшего расселения. В географическом масштабе следствие такой «разборчивости» иллюстрирует распространение дождевых червей в Северной Америке, где его связывают с висконсинским оледенением, закончившимся более 10 тыс. лет назад. Будучи истреблены Канадским ледниковым щитом, имевшим мощность до трех километров и занимавшим почти всю Канаду и частично США, черви не вернулись на некогда занимаемые территории. Северная граница их распространения проходит до сих пор в среднем по южной границе ледникового щита [3]. Эта устоявшаяся точка зрения между тем нуждается в доказательстве, поскольку причина может заключаться не только в палеогеографической истории и «домоседстве», но и в современных значениях зимних температур, которые ограничивают распространение червей на север.
![Распространение аборигенных (слева) и «экзотических» для Северной Америки (завезенных) дождевых червей ([3] с изменениями). Жирные линии — южная и северная границы ледника во время последнего ледникового максимума](/sites/default/files/inline-images/1465_3.jpg)
Территориальный консерватизм и отсутствие «перемешивания» генофонда подразумевает слабый обмен генами между популяциями, слагающими вид, что ведет в конце концов к накоплению достаточных различий, предотвращающих скрещивание или делающих потомство нежизнеспособным, т.е. к дивергенции. Иными словами, применительно к дождевым червям (не космополитам!) географический барьер как необходимое условие одного из способов разделения популяций и последующего видообразования может вырождаться во внешне незаметное и трудно выявляемое препятствие.
Естественно ожидать, что в пределах одного вида, выделенного по морфологическим критериям и имеющего значительный ареал, генетические различия могут (или даже — должны) быть высоки.
ДНК-штрихкодирование
Действительно, изменчивость ДНК большинства видов дождевых червей весьма значительна. Одна из первых работ, проведенных методами ДНК-штрихкодирования [4], выявила в пределах нескольких широко распространенных видов от двух до пяти криптических генетических линий. Они заметно отличались друг от друга: доля попарных замен между линиями достигала 22%. Подобная картина обнаружена и в последующих экспериментах, как на отдельных видах, так и на обширных их рядах.
Почти «чистое поле», которому следует уподобить степень изученности генетической изменчивости дождевых червей равнин России, и желание при минимальных затратах получить хотя бы контуры ожидаемой картины определяют применение метода ДНК-штрихкодирования. Его основной принцип — использование коротких фрагментов генома для идентификации видов [5]. Наиболее часто используемая для ДНК-штрихкодирования последовательность у животных — митохондриальный ген цитохромоксидазы 1 (cox1). Этот метод очень полезен при исследовании образцов, которые не могут быть достоверно определены традиционными способами, например ювенильных особей, коконов, фрагментов организмов или следов их жизнедеятельности (фекалий, содержимого желудка и т.д.), а также ДНК, выделенной из образцов почвы. Применяется он и для групп с большим числом трудно определяемых видов или для таких, систематика которых плохо разработана. ДНК-штрихкодирование имеет более ограниченный круг задач, чем молекулярная филогения. В частности, они не включают исследование таксономических (родственных) взаимоотношений между видами или уточнение их систематического положения. Тем не менее метод в некоторых случаях позволяет находить новые виды и приблизительно устанавливать их родство. Хотя само по себе ДНК-штрихкодирование не может лежать в основе таксономических исследований, оно позволяет идентифицировать предполагаемые новые виды.
Перспективные образцы уже исследуются с помощью маркеров ядерной ДНК. Пока мы ограничиваемся транскрибируемыми спейсерами рибосомального кластера, которые могут подтвердить или не подтвердить высокий уровень различий, выявленный ДНК-штрихкодированием. Для выяснения же точных родственных взаимоотношений между генетическими линиями или видами потребуется анализ целого набора ядерных генов.
Дождевой червь Eisenia nordenskioldi — замечательный пример вида со столь обширным ареалом, что генный обмен (уж во всяком случае между краевыми популяциями) за срок со стабильной по палеогеографическим меркам обстановкой можно полностью исключить. В совсем недавнем прошлом он представлялся как единый, морфологически (кроме пигментации) неделимый вид с двумя подвидами — E. n. nordenskioldi и E. n. pallida. Подвиды различаются распространением, пигментацией, плоидностью и относятся к разным морфоэкологическим группам. Номинативный подвид — единственный дождевой червь, естественный ареал которого лежит в тундровой, лесной и лесостепной зонах, занимая в их пределах почти всю азиатскую и часть восточноевропейской территории России [7]. Он населяет самые суровые регионы Северной Азии. Описан в 1879 г. и назван в честь выдающегося полярного исследователя. Если бы червь не получил имя А. Норденшельда, ему можно было бы, опираясь на ареал, присвоить титул «арктический», «полярный», «северный» и т.п.
Трудно представить, чтобы такое гигантское пространство занимал один вид, к тому же (как считалось) с более чем скромными возможностями к расселению, а значит — с ограниченным генным обменом.
Предположение оправдалось в самой большой степени! Молекулярно-генетические исследования выявили весьма сложную, но ожидаемую картину. Как оказалось, номинативный подвид E. n. nordenskioldi есть не что иное, как комплекс выделенных девяти (на данный момент!) генетических линий.
Второй подвид, E. n. pallida, немногим уступил номинативному: он представляет собой «пучок» из пяти генетических линий того же масштаба, что и в пределах E. n. nordenskioldi. И тут правильнее сказать: «пока из пяти». Но о комплексе E. n. pallida — специальный подробный разговор в другой раз.
В целом же очевидно, что E. nordenskioldi — «сборный» таксон, объединяющий много морфологически неразличимых видов; их как минимум 14 (9 + 5).
Ареалы генетических линий
Важнейшая черта обнаруженных линий (номера им присваивались по ходу выявления) номинативного подвида E. nordenskioldi, в еще большей мере подчеркивающая их индивидуальность, — уникальность абрисов и расположения ареалов. Схема сугубо предварительная, так как работа только началась и быстро собрать спиртовую коллекцию с гигантской территории России реальными силами и в обозримый срок в принципе невозможно. Поясним: музейные коллекции дождевых червей в большинстве своем хранятся в убийце ДНК — формалине, не обесцвечивающем тканей. Так что все надо начинать с нуля... Тут самое место поблагодарить коллег и просто отзывчивых знакомых, бескорыстно помогающих нам. Особая благодарность — коллегам из Института проблем экологии и эволюции имени А. Н. Северцова РАН (Москва) и Института биологических проблем криолитозоны СО РАН (Якутск), весьма тщательно собравшим коллекции червей, которые сейчас находятся в обработке.
К настоящему времени наиболее полные картины получены по юго-востоку европейской части России и Южному Уралу, отдельным регионам юга Сибири и Дальнему Востоку. Плохо пока обследованы европейская часть России (кроме уже названной территории), большая часть Западной Сибири (кроме ее юга); совершенно нетронуты Восточная Сибирь и другие регионы. Досадно, что руки пока не дошли до средней и северной тайги востока Западной Сибири и Восточной Сибири, поэтому нельзя обсуждать, вероятно, важнейшую для Сибири первую линию E. n. nordenskioldi, которая, судя по всему, населяет большую часть ее территории. Сборы червей из долины Енисея мало что дадут для искомой картины, так как там нет вечной мерзлоты и, вероятно, будут превалировать в основном завезенные виды. Но ряд нетривиальных заключений, обнадеживающих перспективой, сделать уже можно.
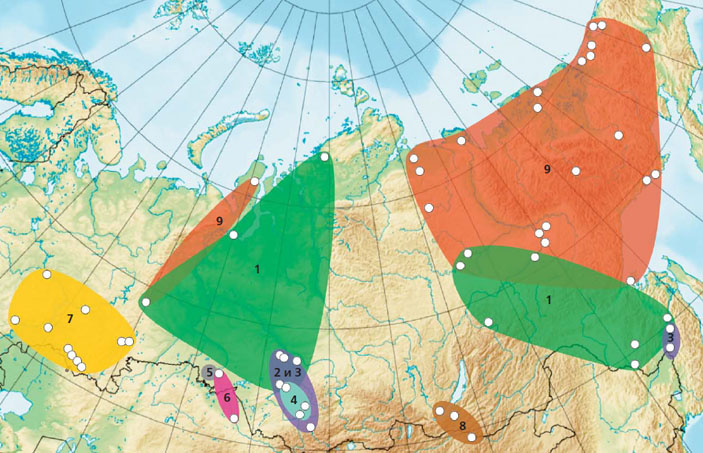
На юго-востоке европейской части России и на Южном Урале, несмотря на значительное число точек, откуда взяты образцы червей, обнаружены E. n. nordenskioldi исключительно седьмой линии. Замечательно, что к востоку от выявленного ареала черви этого кластера не найдены. Но они могут быть северо-западнее (по дубравам) вплоть до Оки в районе Серпухова, если там, конечно, не обитает какая-либо еще не описанная линия.
Как давно обособились европейско-уральские популяции от сибирских линий вида? При всех недостатках метода молекулярных часов он пока единственно возможный и доступный. Учитывая же, что получаемые в результате времена измеряются многими сотнями тысяч лет, можно оперировать лишь порядками величин, переводя их в качественные оценки типа «до того..., после того...», имея в виду крупные подразделения геологического времени. Так вот, черви седьмой линии обособились от сибирских линий свыше миллиона лет назад, т.е. в любом случае в плейстоцене.
В пределах этой линии выделяются две группы: первая — европейские популяции и черви из двух точек с западных склонов юга Южного Урала, вторая — остальные уральские гаплотипы. Судя по топологии филогенетического дерева, европейские популяции вида происходят от одной из ветвей второй группы. Возраст европейской группы — не менее 500 тыс. лет, так что E. n. nordenskioldi проник в Европу давно и пережил там несколько криохронов. Выявленная ныне область распространения червей седьмой линии лежит за пределами даже Днепровского оледенения (средний плейстоцен, 300–250 тыс. лет назад). Упомянутые выше самые северо-западные находки червя (на Оке), возможно, позволят связать его распространение с более поздним — Московским оледенением (средний плейстоцен, около 170–125 тыс. лет назад).
На северо-востоке России выявлена только одна — девятая — линия E. n. nordenskioldi. Таким образом повторяется ситуация, описанная на юго-востоке европейской части России и на Южном Урале. Относящиеся к девятой линии гаплотипы обнаружены также на севере Сибири, в Якутии и на юго-востоке Северного Урала (окрестности г. Карпинска), что указывает на широкое распространение червей этой линии в прошлом.
Каждая из тестированных популяций девятой линии уникальна по генетическому разнообразию, и анализ молекулярных часов свидетельствует, во-первых, об их давней дивергенции, вероятно, в начале плейстоцена, во-вторых — об упомянутой территориальной консервативности («оседлости») червей.
Причины невысокой скорости их расселения на северо-востоке Азии конкретны и потому, как кажется, понятны. Вероятно, важнейший фактор, ограничивающий ландшафтное и биотопическое распределение червей E. n. nordenskioldi по крайней мере девятой линии, — кислотность почвы (pH < 5). В бассейне верховий Колымы доминирующая горная порода (она же — материнская для почв) — разного типа сланцы, содержащие мало карбонатов, что в конце концов сказывается на низких значениях pH почвы и воды. На северных склонах водный pH в почвах очень низок (до 3,5), под разнотравными парковыми лиственничниками — поднимается до 4,5. Лишь в редких осинниках по окраинам степных участков, на горно-тундровых луговинах, в дриадниках и кое-где еще pH заметно выше, достигает 6. Но все эти группировки представляют собой всегда единичные вкрапления в убогий ландшафт лиственничных редколесий, к тому же они крошечные по площади и, конечно, никак не соединяются между собой. Но именно к ним и приурочены микропопуляции дождевых червей. Неслучайно даже «полевой народ» (геологи, охотники, рыбаки и т.д.), как правило, не знает о существовании дождевых червей в континентальных регионах северо-востока Азии. Сколько длится одинокое житие таких «заимок», сказать трудно (нужна статистическая оценка изменчивости), но ясно, что очень долго.
Расселение, очевидно, может быть и относительно быстрым — за счет переноса животных водотоками. В воде подходящей кислотности черви могут жить неопределенно долго, поэтому перенос их таким путем на большие расстояния вполне возможен. Вероятно, поэтому черви из окрестностей пос. Сеймчан и из устья Колымы, находящегося более чем в 1,5 тыс. км ниже, относятся к одному гаплотипу. Это не означает, что конкретный червь доплывает сверху донизу Колымы. Видимо, путь разбит на дистанции: червей можно найти лишь кое-где на крупных участках ивняков в пойме реки. Но к колымскому кластеру принадлежат и черви с южного побережья Чаунской губы (о. Айопечан), которая не только не связана водными артериями с Колымой, но и находится далеко от нее. С другой стороны, черви из окрестностей Певека, что всего лишь в сотне километров от названного острова, весьма далеки генетически, хотя никаких очевидных преград (кроме низких пологих увалов с горными тундрами) на их пути нет. По приморским равнинным тундрам червь E. n. nordenskioldi девятой линии мог расселяться относительно свободно. Благодаря морским (высококарбонатным) отложениям pH почв достигает 7 и таким образом не препятствует существованию червей.
Судя по распространению и возрасту популяций, девятая линия пережила на северо-востоке Азии несколько циклов похолодания—потепления и связанные с ними радикальные ландшафтные изменения. Сказанное неудивительно, так как животные этой линии обладают чрезвычайно широкой экологической валентностью в отношении температуры и влажности. Они — рекордсмены холодоустойчивости: переносят до ?35°С, а откладываемые ими яйцевые коконы — и того больше, до ?45°С. Даже на полюсе холода, в Оймяконской котловине Якутии, температуры в почве на глубине зимовки червей (10–20 см) не бывают столь низки. Но в ледниковое время такая устойчивость была весьма востребована. Черви девятой линии переносят и сухость, точнее — настоящую засуху: они могут находиться в воздушно-сухой почве более двух месяцев, устраивая из кожных выделений и пылеватых частиц почвы защитную капсулу. Ни какой другой вид дождевых червей столь редким сочетанием способностей не обладает.
Подчеркнем, что речь идет о девятой линии, а обладают ли другие линии E. n. nordenskioldi подобными свойствами — большой вопрос, нуждающийся в специальной проработке. Судя по ареалу девятой линии, скорее всего, — нет, не обладают.
Обнаружение червей девятой линии на северо-востоке Азии, несомненно, соответствует представлениям геоморфологов и палеогеографов об отсутствии покровных оледенений в регионе. Ледники здесь были лишь горно-долинными, что создало предпосылки для сохранности на огромном пространстве от верховий Яны и Индигирки до Чукотского п-ова реликтов разного происхождения: тундростепных, степных, неморальных, темнохвойно-таежных. Они свидетельствуют о преемственности в течение длительного времени, возможно, с раннего плейстоцена, ландшафтов (тундростепей) с одновременным присутствием степных и тундровых видов животных и растений. О былом сочетании, казалось бы, несочетаемого можно судить по многочисленным останкам животных, захороненным главным образом в вечной мерзлоте Якутии, северо-востока Азии и северо-запада Америки.
Но останков обитателей доплейстоценовых ландшафтов сохранилось немного, и потому судить об обстановке тех времен сложно. Любое новое свидетельство, пусть и косвенное, — ценность.
Вопреки чрезвычайно широкой экологической валентности девятой линии E. n. nordenskioldi и множественности эпизодов осушения Берингова пролива червь в Восточной Берингии (т.е. в Северной Америке) неизвестен. Заметим, что, по оценкам метода молекулярных часов, дивергенция чукотских популяций девятой линии произошла более 100 тыс. лет назад, задолго до осушения Берингова пролива в период последнего ледникового максимума. Причина отсутствия червя в Восточной Берингии, возможно, кроется в его территориальной консервативности. Пассивное же его расселение из Азии в Америку (например, водотоками) маловероятно, поскольку основные палеореки на осушенном дне пролива были ориентированы в меридиональном направлении, а не в широтном. Между тем непреднамеренная интродукция в Северной Америке червей первой, третьей и девятой линий представляется весьма вероятной, что не должно привести к каким-либо очевидным нежелательным последствиям.
Изложенные предварительные результаты позволяют считать перспективным обследование ареала E. nordenskioldi, особенно северных территорий, по более частой сетке, чтобы уточнить распространение выявленных линий в необследованных еще регионах. Изучение филогеографии обсуждаемого вида и, возможно, других представителей почвенной фауны, переживших, в отличие от мамонта, шерстистого носорога и других животных мамонтовой фауны, плейстоцен, может внести нетривиальный вклад в реконструкцию истории биоты Берингии.
С юго-востоком европейской части и северо-востоком России нам несказанно повезло: как было показано, и там и там обитают черви линий, нигде более не встречающихся (произнесем как заклинание — «пока не найденных»). С другими регионами все не так просто. Генетическое разнообразие E. n. nordenskioldi, широко распространенного в Западной и Средней Сибири (к северу от Транссиба), пока обследовано лишь в немногих точках этого региона. В южных частях всей Сибири и Дальнего Востока собран весьма обширный материал, но его предварительная обработка дала, увы, необнадеживающий результат: пока встречены семь линий из девяти уже выявленных. В большинстве регионов (кроме Забайкалья) линии сосуществуют попарно, а в северных отрогах Алтая, в частности на Салаирском кряже, на юге Новосибирской и Томской областей, обнаружены даже четыре линии. Недостаточно и точек обследования — нужна более частая сетка, нужны и большего размера выборки, чтобы видеть изменчивость.
Сложность изучения Южной Сибири и Дальнего Востока еще и в том, что, в отличие от средней и тем более северной тайги, здесь обитает не только пресловутый E. n. nordenskioldi, но и второй подвид — E. n. pallida, а также много других видов дождевых червей, и не только космополитов.
Юг Западной Сибири мы избрали в качестве полигона для оценки возможного занижения генетического разнообразия дождевых червей. В 12 точках Новосибирской, Томской, Кемеровской областей, в Алтайском крае и Республике Алтай были собраны 250 червей. По данным Кадастра дождевых червей [2], в России и на сопредельных территориях обитает 16 видов. Все они принадлежат к семейству Lumbricidae. Восемь из них — инвазивные виды-космополиты (Allolobophora parva, Dendrodrilus rubidus tenuis, D. r. subrubicundus, Octolasion tyrtaeum (или O. lacteum), Aporrectodea caliginosa, Dendrobaena octaedra, Eiseniella tetraedra и Eisenia fetida). Для Западной Сибири указаны также Lumbricus terrestris, L. castaneus, L. rubellus и Aporrectodea rosea [8]. Четыре вида рода Eisenia (E. malevici, E. altaica, E. salairica и E. tracta) — эндемики Алтая и Салаирского кряжа. Остальные пять — это близкородственная группа Eisenia n. nordenskioldi / E. n. pallida / E. atlavinyteae, а также менее распространенные E. sibirica и E. balatonica. E. atlavinyteae нигде далее не упоминается по весьма прозаической причине: не удается сопоставить этот тяжело определяемый таксон с выделяемыми линиями.
Прежде чем переходить к результатам анализа, необходимо сделать техническое замечание. Известно, что на построенных по последовательностям оснований митохондриальной ДНК деревьях, как правило, не выделяются ветви, соответствующие таксонам надвидового ранга, что обусловлено высокой скоростью эволюции митохондриальной ДНК у дождевых червей. Многие роды и даже семейства на таких деревьях оказываются полифилетичными. В связи с этим кластеризацию проводили следующим образом: кластером, соответствующим одной операционной таксономической единице (Operational Taxonomic Unit, OTU), считали ветвь с бутстрепной поддержкой не менее 95, которая не объединяется с какой-либо другой ветвью на дереве с бутстрепной поддержкой более 90.
Таким путем в выборке были выделены 27 кластеров. Семь из них относятся к широко распространенным видам (A. caliginosa, E. fetida, E. balatonica, E. sibirica, O. tyrtaeum, D. r. tenuis, D. octaedra). Три известных в Западной Сибири вида-космополита (E. tetraedra, D. r. subrubicundus, A. parva), а также алтайские эндемики отсутствуют — очевидно, в связи с тем, что характерные для них биотопы не были обследованы. Три кластера соответствовали генетическим линиям E. n. nordenskioldi [13]. Оставшиеся 17 из 27 кластеров не имели близкого сходства ни с одной из ранее изученных последовательностей ДНК дождевых червей. Девять кластеров из 17 были сходны с группой E. n. nordenskioldi / E. n. pallida.
Новые виды?
Можно ли считать, что выявленные кластеры действительно соответствуют видам дождевых червей? Для кольчатых червей характерен более высокий уровень дивергенции последовательностей мтДНК между видами одного рода по сравнению с другими животными [5]. По данным авторов этой статьи, степень внутривидовой изменчивости у животных редко превышает 2%. Однако уровни попарной дивергенции между генетическими линиями E. nordenskioldi составляют от 16 до 29%. Генетическая изменчивость внутри линий этого вида, для которых имеются данные по достаточному количеству популяций, превышает 5% и доходит до 8,6% у второй линии E. n. pallida. Таким образом, если судить по степени изменчивости мтДНК, то обнаруженные нами кластеры можно уверенно считать отдельными видами.
Один из характерных участков смешанных лесов в долине Амура и на Северном Сихотэ-Алине (Природа» №5, 2016)
Да, бывают случаи, когда представители значительно различающихся митохондриальных генетических линий одного вида могут не иметь существенных различий по ядерным маркерам, что показано, например, для видов Allolobophora chlorotica [4, 14], L. rubellus [15], а также для второй и третьей линий E. n. pallida (наши неопубликованные данные). Причины такого несовпадения пока не объяснены, однако понятно, что существование предполагаемого нового вида дождевых червей необходимо подтверждать и с помощью ядерных маркеров. Тем не менее в большинстве описанных случаев расхождений между генетическими линиями по ядерным и митохондриальным маркерам нет. Их отсутствие свидетельствует о том, что обнаруженные нами кластеры главным образом — новые виды дождевых червей.
Как видно, даже весьма ограниченная выборка (напомним, всего-то 250 червей из 12 точек) из Западной Сибири позволила выявить 17 новых генетических кластеров дождевых червей. Значительная их часть соответствует новым таксонам, что как минимум удваивает ранее известное на исследованной территории число видов.
***
Заключение, следующее из всего изложенного, и радует и тревожит. Радует тем, что проведенная работа подтверждает предположения о высокой криптической изменчивости дождевых червей равнин России и свидетельствует о перспективности исследований в этом направлении. Похоже, не любой дождевой червь из неосвоенных регионов Сибири будет, как прежде, не глядя отнесен к отслужившему свое виду E. nordenskioldi. Появляется некоторая надежда на использование результатов для палеогеографических реконструкций, может быть, даже доплейстоценовых. Тревожит предстоящей грандиозной таксономической работой по описанию новых видов и доскональной ревизией ранее описанных. Многое заново! Дождевые черви столь значимы в природе, что их надо знать персонально и в деталях.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект 10-04-00425а и 16-04-00082а).
Литература
1. Hendrix P. F., Callaham M. A., Drake J. M. et al. Pandora’s box contained bait: the global problem of introduced earthworms // Ann. Rev. Ecol. Evol. Syst. 2008. V. 39. P. 593–613. DOI: 10.1146/annurev.ecolsys.39.110707.173426.
2. Всеволодова-Перель Т. С. Дождевые черви России: Кадастр и определитель. М., 1997.
3. Reynolds J. W. The distribution of earthworms (Annelida, Oligochaeta) in North America // Advances in ecology and environmental sciences / Eds P. C. Mishra, N. Behera, B. K. Senapati, B. C. Guru. New Delhi, 1995. P. 133–153.
4. King R. A., Tibble A. L., Symondson W. O. C. Opening a can of worms: unprecedented sympatric cryptic diversity within British lumbricid earthworms // Mol. Ecol. 2008. V. 17. P. 4684–4698. DOI: 10.1111/j.1365-294X.2008.03931.x.
5. Hebert P. D. N., Ratnasingham S., Waard J. R. de. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species // Proc. R. Soc. Lond. B. 2003. V. 270. P. 96–99. DOI: 10.1098/rsbl.2003.0025.
6. Шеховцов С. В., Берман Д. И., Пельтек С. Е. Филогеография дождевого червя Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) на северо-востоке Евразии // ДАН. 2015. Т. 461. № 1. С. 118–121.
7. Перель Т. С. Распространение и закономерности распределения дождевых червей фауны СССР. М., 1979.
8. Стриганова Б. Р., Порядина Н. М. Животное население почв бореальных лесов Западно-Сибирской равнины. М., 2005.
9. Всеволодова-Перель Т. С. Распространение дождевых червей на севере Палеарктики (в пределах СССР) // Биология почв Северной Европы / Ред. Д. А. Криволуцкий. М., 1988. С. 84–99.
10. Всеволодова-Перель Т. С., Лейрих А. Н. Распространение и экология дождевого червя Eisenia nordenskioldi pallida (Oligochaeta, Lumbricidae), массового на юге Сибири и Дальнего Востока // Зоол. журн. 2014. Т. 93. № 1. С. 45–52.
11. Берман Д. И., Мещерякова Е. Н. Ареалы и холодоустойчивость двух подвидов дождевого червя (Eisenia nordenskioldi, Lumbricidae, Oligochaeta) // Зоол. журн. 2013. Т. 92. № 7. С. 771–780.
12. Ганин Г. Н. Почвенные животные Уссурийского края. Владивосток-Хабаровск, 1997.
13. Shekhovtsov S. V., Golovanova E. V., Peltek S. E. Mitochondrial DNA variation in Eisenia n. nordenskioldi (Lumbricidae) in Europe and Southern Urals // Mitochondrial DNA. 2015. DOI: 10.3109/19401736.2015.1101594.
14. Dupont L., Lazrek F., Porco D. et al. New insight into the genetic structure of the Allolobophora chlorotica aggregate in Europe using microsatellite and mitochondrial data // Pedobiologia. 2011. V. 54. P. 217–224. DOI: 10.1016/j.pedobi.2011.03.004.
15. Giska I., Sechi P., Babik W. Deeply divergent sympatric mitochondrial lineages of the earthworm Lumbricus rubellus are not reproductively isolated // BMC Evolutionary Biology. 2015. V. 15. P. 217–229. DOI: 10.1186/s12862-015-0488-9.